
Aquí se revisa el conocimiento actual sobre las consecuencias de la invasión de Acacia en áreas mediterráneas, así como se plantean ideas para el control y recuperación de áreas invadidas y se sugieren perspectivas futuras de investigación.
Presentación
“Here to stay. Recent advances and perspectives about Acacia invasion in Mediterranean areas” (Aquí para quedarse. Avances recientes y perspectivas sobre la invasión de Acacia en áreas mediterráneas”) es un artículo tipo review publicado en la revista Annals of Forest Sicence por Souza-Alonso, P., Rodríguez, J., González, L. y Lorenzo, P., de las Universidades de Vigo y de Coimbra.
En él, los autores revisan el conocimiento actual sobre las consecuencias de la invasión de Acacia en áreas mediterráneas. También recogen y proponen diferentes ideas para el manejo y recuperación de áreas invadidas, así como sugieren perspectivas futuras para la investigación sobre el género Acacia.
Para ello, han recompilado, resumido y discutido recientes descubrimientos sobre aspectos físico-químicos, ecológicos, microbiológicos y socioeconómicos de la invasión de Acacias australianas (Acacia dealbata, A. longifolia, A. mearnsii, A. saligna y A. melanoxylon) centrándose en áreas mediterráneas.
Tras esto, se ha visto que la invasión de Acacia conlleva generalmente alteraciones físico-químicas del suelo y cambios en la función y estructura microbiana. Consecuencias como la disminución de biodiversidad, alteración de la estructura del ecosistema, bancos de semillas más grandes dominados por especies de plantas invasoras, nuevas relaciones biotróficas o alteraciones en la disponibilidad de agua y el régimen de incendios; sugieren que las Acacias están incluso creando nuevos ecosistemas a nivel local.
Por ello, se concluye que tanto la previsión de invasiones, como la modelización y el manejo de ecosistemas dominados por acacias, son tareas desafiantes que deben ser corregidas en el futuro, ya que las condiciones climáticas y la intensificación de los usos del suelo están aumentando la probabilidad de que haya una mayor invasión de Acacia en áreas mediterráneas. Además, las acciones de manejo llevadas a cabo sin éxito, sugieren que la restauración debería ser meticulosamente monitorizada, pero la magnitud de la invasión o la inconsistencia de la inversión económica indican que la erradicación es a menudo inviable.
Alternativamente, son necesarias nuevas soluciones integrativas y económicas, incluyendo la colaboración de la sociedad, políticos y partes interesadas, para prevenir nuevas introducciones y alcanzar un control sostenible de las acacias. Además, hay que indicar que actualmente hay un creciente interés en investigación aplicada a la valorización de nuevos usos de las Acacias y sus residuos, que da lugar también a beneficios económicos.

Se sabe que las plantas exóticas invasoras (IAPs) alteran fuertemente los ecosistemas reduciendo la biodiversidad, modificando los servicios de dicho ecosistema y aumentando los impactos negativos en el nivel socio-económico. Entre los taxones invasores, la Acacia es un género altamente problemático en todo el mundo. De hecho, casi 500 artículos han sido publicados sobre diversos aspectos de invasiones de Acacia en los últimos 25 años.
Existen 24 especies del género Acacia confirmadas como invasoras en todo el mundo. En este trabajo se han centrado en el subgénero Phyllodineae Acacia s.s. por su invasividad, tratando las siguientes especies: Acacia dealbata Link, A. longifolia (Andrews) Willd, A. mearnsii De Wild., A. saligna (Labill) H. L. Wendl. y A. melanoxylon R. Br., por sus impactos en todo el mundo.
El movimiento de acacias hacia otros continentes comenzó a finales del 1700, sin embargo, en los últimos 200 años, han tenido lugar velocidades de dispersión sin precedentes, siendo ahora frecuente encontrar invasiones de acacias australianas en áreas mediterráneas. La siguiente Figura 1 representa el número de artículos (papers) que incluyen estas acacias altamente invasivas a lo largo de todo el mundo.
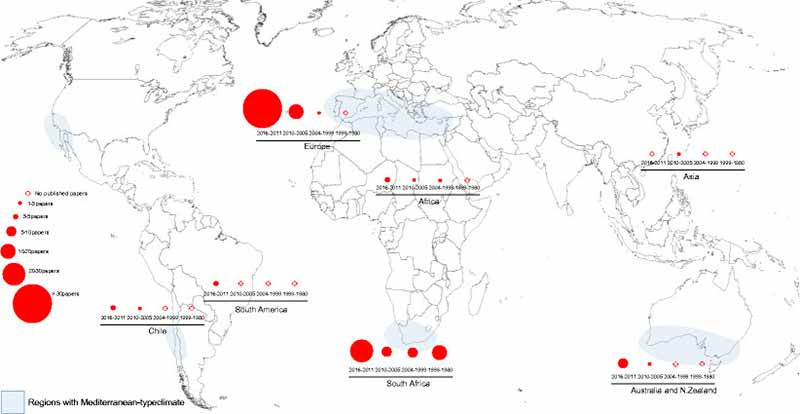
Características específicas de estas especies de acacias, como adaptabilidad a muchas condiciones medioambientales, fácil germinación y crecimiento, buena supervivencia y tasas de crecimiento rápido, valor ornamental o la calidad de su madera, han determinado su distribución actual. Además, en Europa y áreas mediterráneas, algunos usos de las acacias como la producción de madera, en la industria del perfume, en estabilización de dunas y para evitar la erosión de la arena o estabilizar rampas y colinas derivadas de la construcción de vías del tren; tienen un importante papel en su introducción y como consecuencia, ahora muchos biomas mediterráneos se encuentran amenazados por las acacias.
De hecho, las acacias han sido denominadas como “transformers”, ya que son especies que “sustancialmente cambian la personalidad, condición, forma o naturaleza de los ecosistemas, convirtiéndose en agentes activos en procesos de formación de regiones” (Richardson et al. 2000a; Marchante et al. 2011a). Sin embargo, a pesar de esto, la percepción humana de la Acacia como invasora es muy variable según las circunstancias, ya que en muchos sitios la Acacia dealbata está bien vista como fuente de material de construcción, de medicina o calor, por lo que puede ser cultivada para un beneficio económico e incluso se llevan a cabo celebraciones en su honor, pero si ésta afecta a cultivos negativamente, es reconocida como planta problemática.
Métodos
Como se ha comentado, los autores de este trabajo han recompilado, resumido y discutido recientes descubrimientos sobre aspectos físico-químicos, ecológicos, microbiológicos y socioeconómicos de la invasión de acacias australianas en áreas mediterráneas. A continuación, se habla un poco de cada uno.
Genética, fenotipo y fisiología
Se sugirió que el tamaño del conjunto de genoma podría ser un factor que promoviera la invasión, sin embargo, hay estudios que demuestran que este hecho es falso, ya que no se hallaron diferencias en el tamaño del genoma basado en el carácter invasivo entre acacias invasivas y no invasivas cuando éstas eran introducidas en otro ecosistema. Además, se vio que niveles bajos de diversidad genética en las áreas introducidas comparados con áreas nativas, no están relacionado con el éxito de invasión.
Por su parte, los rasgos ecofisiológicos pueden ser tan importantes como los morfológicos para explicar la invasividad. Una vez que las semillas llegan al suelo, las acacias están dotadas de mecanismos para superar a las especies nativas. Por ejemplo, en ambientes con salinidad hídrica, las semillas y las plántulas de A. longifolia presentan concentraciones aumentadas de iones intracelulares, absorción eficiente de nitrógeno, defensa contra radicales superóxido y alta tolerancia a una amplia gama de concentraciones de sal, en comparación con especies nativas (Morais et al., 2012 Morais y Freitas 2012).
Otro ejemplo es que A. melanoxylon se comporta mejor cuando la temperatura de la hoja es más alta que la de las acacias nativas bajo estrés hídrico. Este hecho podría reflejar una mayor termoestabilidad del PSII (fotosistema 2) o, por el contrario, una mejor aclimatación y, por tanto, lograr un eficiente proceso fotosintético en ecosistemas áridos o de tipo mediterráneo. Además, bajo condiciones experimentales, se ha relacionado un aumento de CO2 con mayores tasas de crecimiento, peso final y mayores tasas de fijación de N de A. melanoxylon; por consiguiente, si la disponibilidad de N aumenta, la biomasa seca, la asimilación de CO2, el espesor del follaje y la densidad aumentan también significativamente (Schortemeyer et al., 1999).

Así pues, en un esperado escenario de calentamiento global con mayores temperaturas y niveles de CO2, con las acacias creciendo a tasas más altas y produciendo capiteles con follaje más denso, reduciendo la disponibilidad de luz para especies de sotobosque, la invasividad de estas especies podría verse severamente incrementada.
Características reproductivas
Se sabe que la reproducción por brote facilita el establecimiento de poblaciones clonales. De hecho, la proporción de especies germinantes es mayor entre las acacias invasivas que las no invasivas. Las acacias invasoras también alcanzan la madurez reproductiva antes (<2 años) que las no invasoras (Gibson et al., 2011). Acacia dealbata y A. mearnsii tienden a tener niveles más altos de autocompatibilidad, lo que sugiere que la habilidad de autofertilizarse puede favorecer su invasividad. Por su parte, A. saligna tiene un sistema de apareamiento mixto, cruzamiento preferencial, pero también con cierto nivel de autofecundación (George et al., 2008).
Hay que indicar que la autopolinización podría ser una valiosa herramienta para producir descendencia en circunstancias que restringen severamente la supervivencia de las plantas (por ejemplo, áreas aisladas, ausencia de polinizadores o limitación de los compañeros). Curiosamente, A. mearnsii mostró una reproducción sexual y asexual dependiendo de las condiciones ambientales en el rango no nativo, mostrando preferencia por el brote en áreas perturbadas y la reproducción basada en semillas en sitios no perturbados (Eilu y Obua 2005).

A pesar de su capacidad de autofertilización, las acacias son polinizadas por insectos generalistas y por lo general, requieren la presencia de vectores de polinización para lograr una producción de semilla significativa (Correia et al., 2014). El éxito reproductivo a menudo se maximiza por la apertura sincronizada y masiva de las flores, tanto dentro de un solo individuo como de las poblaciones locales, lo que puede interferir con la relación normal entre especies nativas y sus polinizadores.
En cuanto a las semillas, se sugiere su producción como factor promotor de la invasión de Acacia en áreas mediterráneas (Castro-Díez et al., 2011). Así pues, en el área introducida, A. dealbata y A. longifolia escapan a la depredación y exhiben una mayor producción de semillas completamente desarrolladas por fruto (A. longifolia) o por árbol (A. dealbata), acompañadas de un tamaño mayor de las semillas individuales.
Después, una vez liberadas, las semillas pueden ser dispersadas por el agua o el viento, pero también a través de animales. Hay que destacar que el fuego estimula la germinación de las semillas de varias acacias invasoras como A. melanoxylon, A. dealbata y A. saligna (García et al. 2007; Lorenzo et al. 2010a; Wilson et al. 2011). Además, la butenolida, un compuesto químico aislado de humo, puede tener un efecto positivo significativo en la ecología de plántulas post-fuego de A. mearnsii (Kulkarni et al., 2007). El efecto estimulante del fuego tiene importantes implicaciones ecológicas, ya que éste puede eliminar las semillas nativas de la capa superficial, favoreciendo la germinación de semillas de acacia resistentes y, por tanto, el éxito de la invasión (Richardson y Kluge, 2008; al., 2014). Esto es particularmente relevante para los ecosistemas mediterráneos que se caracterizan por incendios frecuentes, lo que podría contribuir a explicar el éxito de acacias como A. saligna o A. melanoxylon. Además, en un escenario de cambio climático, se esperan incendios forestales extremos y más frecuentes en estos ecosistemas (IPCC 2013), lo que podría ampliar de manera efectiva el área de distribución de las acacias invasoras.

Asociaciones simbióticas
Al igual que el resto de leguminosas, las acacias son altamente dependientes de asociaciones simbióticas con microbios compatibles. En un nuevo hábitat, el acceso a rizobios compatibles es un factor crítico que condiciona la capacidad invasiva de las leguminosas, ya que los mutualismos juegan un papel clave durante su establecimiento (Parker 2001). Las acacias invasoras pueden asociarse con una amplia variedad de bacterias fijadoras de N y su capacidad invasiva podría estar determinada principalmente por la capacidad de formar nódulos profusamente y con mayor eficiencia que las leguminosas N-fijadoras nativas (Rodríguez-Echeverría et al., 2009, 2010).
Las acacias suelen establecer relaciones simbióticas con el género Bradyrhizobium, más específicamente con Bradyrhizobium japonicum, tanto en los rangos nativos como no nativos (Biroudba et al., 2012; Boudiaf et al., 2014). Sin embargo, también se han reportado interacciones simbióticas con nuevos mutualistas en rangos no nativos para acacias australianas, como Bradyrhizobium y Rhizobium, Mesorhizobium, Ochrobactrum y Ensifer meliloti (Rodríguez-Echeverría et al., 2011; Birnbaum et al., 2012). Sin embargo, A. longifoia, A. dealbata y A. melanoxylon se asocian preferentemente con simbiontes co-introducidos en rangos no nativos (Rodríguez-Echeverría et al., 2011, Lorenzo y Rodríguez-Echeverría, 2015), descartando la promiscuidad simbiótica como rasgo invasivo. De hecho, el análisis genético de bacterias simbióticas de nódulos de raíz de A. saligna procedentes de Portugal indicó que las secuencias obtenidas se agruparon principalmente con secuencias australianas, lo que sugiere la co-introducción de parejas simbióticas (Crisostomo et al.2013). En consecuencia, la rápida expansión y gran capacidad de nodulación de A. longifolia podría ampliar también la población y propagación del asociado Bradyrhizobium exótico (Rodríguez-Echeverría et al., 2009).

También se ha sugerido el establecimiento de una retroalimentación positiva del suelo cuando A. dealbata crece en suelos previamente invadidos. Este hecho, ilustra el riesgo ecológico de la introducción voluntaria e involuntaria de microorganismos exóticos mutualistas en proyectos de reforestación. La invasión por especies de Acacia puede estar favoreciendo una segunda invasión por sus microbios exóticos asociados en el suelo. Como consecuencia, tal interacción sinérgica podría acelerar los impactos en los ecosistemas en las áreas introducidas (hipótesis de colapso invasivo, Simberloff y Von Holle, 1999).
Alelopatía
La liberación de aleloquímicos por plantas invasoras se ha postulado como un factor que influye en el medio ambiente circundante y que favorece la invasión (Inderjit et al., 2011). El fenómeno alelopático ha sido ampliamente estudiado en A. dealbata. En las áreas invadidas, los extractos de A. dealbata que contienen concentraciones naturales o cercanas a las naturales, afectaron a la germinación, al crecimiento de las plántulas, a la tasa fotosintética neta, a la tasa de respiración y a la biomasa de las plantas agrícolas y nativas del sotobosque (Carballeira y Reigosa, 1999; 2012, Aguilera et al., 2015a), así como también a la diversidad funcional de los microbios del suelo (Lorenzo et al., 2013a). Además, los experimentos in vitro con lixiviados naturales obtenidos de plantas adultas de A. dealbata aumentan la longitud de radículas de sus propias plántulas, lo que sugiere una autoestimulación (Lorenzo et al., 2010c).
Sin embargo, el efecto estimulador desapareció cuando las plántulas de A. dealbata se cultivaron en suelos nativos (Lorenzo y Rodríguez-E cheverría 2012). A pesar de la evidencia de alelopatía en condiciones controladas, el efecto alelopático no se detectó a escala de campo, lo que sugiere que la alelopatía tiene un papel insignificante durante el proceso vascular de A. dealbata, al menos en el área europea (Lorenzo et al 2016a Souza- Alonso y otros en revisión). Por tanto, en este caso, son necesarios experimentos que imiten las condiciones naturales para identificar claramente el papel de la alelopatía en el proceso invasivo, ya que de lo contrario, el cuadro alelopático de las acacias permanecerá poco claro e incompleto.
Resultados
En cuanto a los resultados de la invasión de Acacia en áreas mediterráneas se vio que ésta conlleva alteraciones físico-químicas del suelo y cambios en la función y estructura microbiana, tales como disminución de biodiversidad, alteración de la estructura del ecosistema, bancos de semillas más grandes dominados por especies de plantas invasoras, nuevas relaciones biotróficas o alteraciones en la disponibilidad de agua y el régimen de incendios; por lo que la previsión de invasiones, modelización y manejo de ecosistemas dominados por acacias, se trata de una tarea que debe ser corregida en el futuro.
Efectos en los ecosistemas
Las acacias invasoras afectan tanto a los compartimentos superficiales como subterráneos, así como a los servicios ecosistémicos, tales como la formación de suelos, el flujo de agua, el ciclo de los nutrientes, la producción de madera y fibra, y las oportunidades recreativas o educativas que sustentan el bienestar humano (Le Maitre et al.2011). Las principales características de las invasiones de acacia están representadas en la siguiente Figura 2, sin embargo, la invasión de acacias presenta diferencias geográficas a lo largo de las regiones mediterráneas.
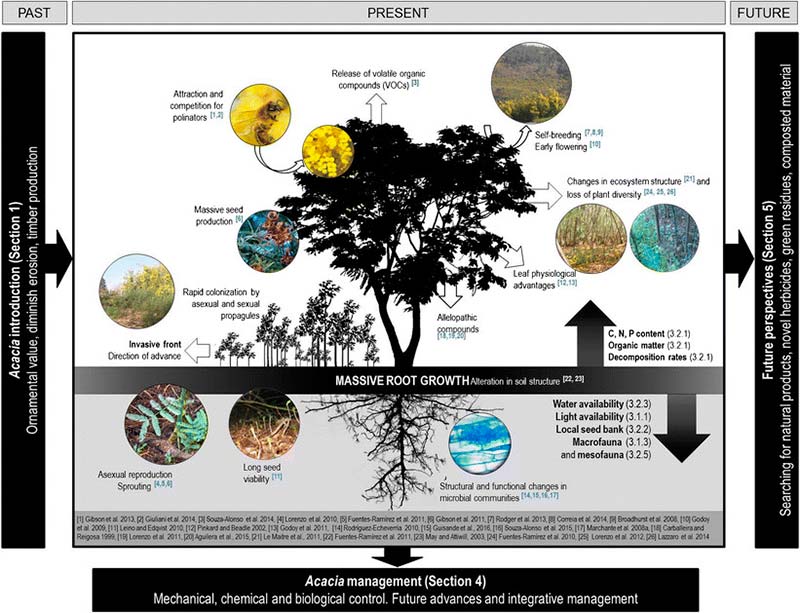
Efectos sobre el suelo:
- Cambios estructurales:
Las acacias invasoras crean formaciones de vegetación homogénea y densa que reducen drásticamente la disponibilidad de luz para las plantas de sotobosque que obstaculizan su establecimiento (Lorenzo et al., 2010a, Lorenzo et al., 2016a). Sin embargo, A. dealbata no reduce la disponibilidad de luz en bosques nativos de hoja ancha (González-Muñoz et al., 2012). Este hecho revela que la influencia de A. dealbata sobre las condiciones de luz es severa en los ecosistemas de doseles abiertos nativos, pero con ligero efecto en los ecosistemas cerrados. Además, los densos acopios conducen a la acumulación de una gran cantidad de biomasa y desechos, lo que aumenta la incidencia y la intensidad de los incendios en los espacios invadidos. Los fuegos, a su vez, estimulan la germinación de las semillas de acacia y reducen la viabilidad de las semillas nativas, favoreciendo así el proceso invasivo (Richardson y Kluge, 2008; Le Maitre et al., 2011).
- Biodiversidad de plantas:
En general, las invasiones de Acacia reducen significativamente la cobertura vegetal, la riqueza de especies y la diversidad. Ésta última, da lugar a la sustitución de especies nativas por otras no nativas o plantas exóticas (Fuentes-Ramírez et al., 2011, Lorenzo y otros, 2012, Marchante et al, 2011b, González-Muñoz et al.)
- Macrofauna:
La presencia de acacias invasoras también modifica la idoneidad del hábitat para los animales y establece nuevas redes ecológicas. Van der Colff et al (2015) encontraron una tendencia diferente de la composición de la comunidad de artrópodos entre áreas nativas e invadidas por A. mearnsii, donde los artrópodos podrían utilizar árboles exóticos como una vía para llegar a hábitats aislados. Además, otros animales más grandes también se ven afectados por las invasiones de Acacia. La densidad de árboles de A. saligna junto factores como la densidad urbana o la estructura de la vegetación, contribuyeron a la disminución de la diversidad de aves (Dures y Cummings 2010) y especies de pequeños mamíferos (Manor et al 2008), vinculando la disminución de la biodiversidad con una reducción en la calidad del hábitat o la integridad del ecosistema. Además, las semillas de A. mearnsii son utilizadas como fuente de nutrientes por el primate especializado Cercopithecus albogularis labiatus, lo que altera su comportamiento alimenticio y probablemente afecta a la propia dispersión de A. mearnsii (Wimberger et al., 2017).

Efectos bajo el suelo:
- Cambios físico-químicos y ciclo de nutrientes:
El abrumador desarrollo de las raíces superficiales de los árboles de Acacia domina y transforma drásticamente la superficie del suelo. Acacia dealbata crea una raíz en la capa superior del suelo debido a su extenso sistema rizomatoso, reduciendo la densidad del suelo (May y Attiwill 2003). Del mismo modo, A. saligna desarrolla raíces alcanzando los 6m durante los primeros 4 años (Knight et al., 2002).
Además, debajo del dosel, una capa gruesa de materia orgánica se acumula progresivamente por la caída continua de la hojarasca (Marchante et al., 2004; Castro-Díez et al., 2012). Las acacias proporcionan desechos con diferentes composiciones de fuentes C que pueden afectar al ciclo de los nutrientes y su descomposición, con posibles ramificaciones ecológicas (Ens et al., 2009a). Sin embargo, el material vegetal descompuesto de A. dealbata no produjo cambios significativos en el perfil funcional y estructural de las comunidades microbianas del suelo y en las propiedades químicas del suelo, en comparación con la descomposición de cantidades similares de material vegetal nativo (Guisande y otros en preparación).
Sin embargo, hay que destacar que como aglomerantes de nitrógeno, las acacias aumentan las concentraciones de N o NH4 + (Castro-Díez et al., 2012). Acacia saligna modifica el ciclo del N a través de la producción de mayores cantidades de desechos, lo que da como resultado que más N se devuelva al suelo y haya un aumento en la disponibilidad de N inorgánico (Yelenik et al., 2004). Acacia longifolia proporciona grandes cantidades de N a la vegetación circundante; sin embargo, al mismo tiempo, requiere cantidades sustanciales de P, lo que crea un desequilibrio N / P a nivel de la comunidad (Ulm et al., 2016). Además, las acacias modifican sustancial y progresivamente el contenido de C en suelos invadidos durante mucho tiempo (Yelenik et al., 2004, Marchante et al., 2008a, Souza-Alonso et al., 2015).


Otros parámetros, como el contenido de materia orgánica o P intercambiable, fueron significativamente incrementados por A. dealbata en suelos de diferentes ecosistemas (Lorenzo et al., 2010b, Souza-Alonso et al., 2014b). Sin embargo, Castro-Díez et al. (2012) no encontraron diferencias en el pH o en la materia orgánica después de la invasión de A. dealbata, por lo que sugirieron que la variación del pH podría ser altamente dependiente del ecosistema estudiado.
Curiosamente, todos estos cambios en el suelo, llevan a una retroalimentación positiva entre las acacias y los suelos invadidos. Los suelos previamente invadidos por A. dealbata favorecen el crecimiento de sus propias plántulas y aumentan la mortalidad de los Pinus pinaster Aiton nativos coexistentes (Lorenzo y Rodríguez-Echeverría 2012; Rodríguez-Echeverría et al., 2013), y este efecto puede continuar incluso después de la eliminación de Acacia (Marchante et al., 2008b, 2011a).
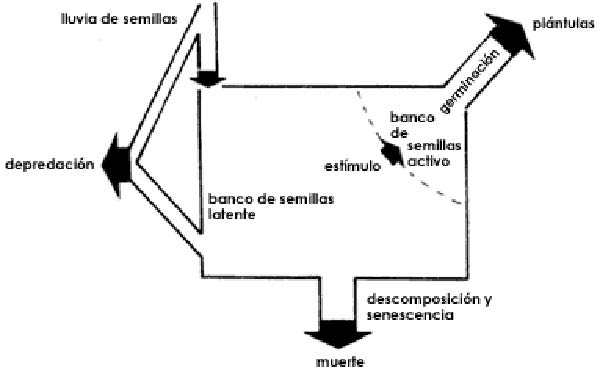
- Banco de semillas:
La composición del banco de semillas del suelo después de la invasión de Acacia se modifica significativamente, limitando o interrumpiendo el suministro de propágulos nativos. Así pues, la riqueza de las semillas nativas disminuyó drásticamente después del aumento de la densidad de A. longifolia, mientras que las semillas del invasor se fueron acumulando progresivamente (Fourie, 2008, Le Maitre et al., 2011). Del mismo modo, la diversidad del banco de semillas en los sábalos invadidos por A. saligna y A. dealbata se vio gravemente afectada, resultando en una disminución y homogeneización en el tamaño de la semilla nativa y porcentajes más altos de semillas exóticas en ecosistemas invadidos (Marchante et al., 2011b, González-Muñoz et al., 2012).

- Relaciones hídricas:
La disponibilidad de agua se indica a menudo como uno de los principales factores limitantes del crecimiento de las plantas en las zonas mediterráneas (Claeys e Inzé 2013, Flexas et al., 2014). Las acacias invasoras son consideradas como especies que consumen agua y su presencia conduce a una reducción en la cantidad y calidad del agua disponible en el suelo y a un aumento en la tasa de evapotranspiración (Lorenzo y Rodríguez-Echeverría 2015). Las poblaciones de A. mearnsii presentaron mayores pérdidas de agua en comparación con los nativos (Dye et al., 2001), mientras que A. longifolia redujo el flujo de agua en un 26% en los bosques de pinos de las dunas costeras de Portugal (Rascher et al., 2011b). Además, hay que indicar que el alto consumo de agua se considera generalmente una estrategia para el crecimiento rápido individual.
Las acacias también pueden influir en la disponibilidad de agua para las comunidades de plantas circundantes a través de otras estrategias a nivel de raíz. Los alcanos de alto peso molecular exudados por las raíces por A. longifolia, pueden inducir la repelencia al agua, reduciendo así el agua accesible para las plántulas nativas (Ens et al., 2009b). Sin embargo, en condiciones estresantes de suministro limitado de agua, A. longifolia reveló una alta sensibilidad a la sequía en términos de biomasa y eficiencia de absorción de N, lo cual fue aún más marcado cuando las plantas crecieron con competencia intraespecífica o interespecífica (Werner et al.2010).
La eliminación de las acacias podría facilitar la reposición de agua para la vegetación nativa, convirtiéndose en un factor clave a considerar en las operaciones de manejo, particularmente en las áreas del Mediterráneo. De hecho, la eliminación de A. mearnsii y A. longifolia de los hábitats ribereños aumentó el caudal (Prinsloo y Scott, 1999). También Jovanovic et al. (2009) indicaron que el desmonte de tierras invadidas por A. saligna, además del aumento en la disponibilidad de agua debido a la reducción de la evapotranspiración, también puede reducir la contaminación de las aguas subterráneas por nitrato.
- Microorganismos del suelo:
Estudios recientes encontraron cambios sustanciales en las comunidades microbianas del suelo a nivel estructural y funcional producidas por la invasión de Acacia (Marchante et al., 2008a, Lorenzo et al., 2010b, Boudiaf et al., 2013, Souza-Alonso et al., 2014b, 2015). Estos cambios son más pronunciados a largo plazo o en áreas fuertemente invadidas y dependen del ecosistema invadido (Marchante et al., 2008a, Lorenzo y Rodríguez Echeverría 2015). Además, las bacterias parecen resultar más afectadas que los hongos (Marchante y otros, 2008a, Lorenzo y Rodríguez-Echeverría, 2015).
- Mesofauna:
Las relaciones entre las plantas nativas y la comunidad de descomponedores también pueden ser alteradas debido a la presencia de acacias. Coetzee et al. (2007) encontraron una reducción significativa en la riqueza, abundancia y tamaño corporal de los artrópodos en pastizales invadidos por A. dealbata en comparación con las áreas no invadidas. Además, la presencia y la producción de desechos de A. mearnsii en los hábitats ribereños alteró la estructura de las comunidades de invertebrados, reduciendo la abundancia de algunos taxones, pero aumentando otros que se alimentan de pequeñas partículas (Lowe et al., 2008). Debajo de los doseles de A. melanoxylon y A. mearnsii, la riqueza de invertebrados se redujo en comparación con aquella bajo especies nativas, y esta reducción fue mayor a nivel de especie que a nivel de familia o de orden (Samways et al., 1996), lo que indica que los cambios en las especies dominantes tiene probablemente implicaciones más bajas a nivel funcional. Además, los cambios cualitativos en la composición de los desechos producidos por la invasión de A. dealbata y A. longifolia, resultan en un pobre material nutritivo para los isópodos terrestres, los cuales son componentes clave de las comunidades de macro descomponedores, lo que conduce a individuos más pequeños (Sousa et al., 1998).
Control y gestión de Acacia en áreas mediterráneas
La investigación sobre la gestión de Acacia comenzó en Sudáfrica, un país pionero que implementa políticas de gestión a nivel nacional. Los primeros esfuerzos organizados para controlar A. dealbata, A. longifolia o A. mearnsii se llevaron a cabo principalmente a través de la implementación del programa “Working for Water” (van Wilgen et al., 2011). En general, la gestión de las acacias es una inversión costosa y una tarea a largo plazo debido a la capacidad de germinación y sus bancos de semillas grandes y resistentes (Richardson y Kluge, 2008; van Wilgen et al., 2016).
Se han logrado resultados efectivos potenciales utilizando el herbicida triclopyr combinado con el corte de individuos de A. dealbata en una estrategia a corto plazo (Campbell y Kluge, 1999; Souza-Alonso et al., 2013). Este herbicida combinado con el corte, fue útil para reducir A. saligna en el control posterior a la quema. Sin embargo, cortar las plantas de A. saligna por debajo del punto de cosecha produjo los mejores resultados (Krupek et al., 2016), con lo que el conocimiento de la mejor etapa fenológica para el manejo de las acacias mejora la efectividad de las acciones de manejo.
Control biológico
El control biológico de las acacias comenzó con la introducción de la avispa, Trichilogaster acaciaelongifoliae, para controlar A. longifolia en Sudáfrica (Dennill y Donnelly 1991). Después de varias generaciones, la producción de vainas de A. longifolia se vio muy reducida. Del mismo modo, el escarabajo Melanterius ventralis se introdujo para alimentarse de semillas de A. longifolia, produciendo mortalidad de semillas en un rango de 15 a 79.5% (Donnelly y Hoffmann 2004). Durante el período 1991-2005, el hongo de roble introducido Uromycladium tepperianum también afectó significativamente a A. saligna al reducir la densidad de árboles (entre 87 y 98%) y la masa del dosel, aumentando también la mortalidad de los árboles (Wood y Morris 2007).

Sin embargo, hay que tener en cuenta que pueden ocurrir efectos secundarios indeseables del control biológico. Por ejemplo, en Sudáfrica, la liberación de algunos agentes de control tales como T. acaciaelongifoliae, Dasineura dielsi y M. ventralis dañó sin intención a A. melanoxylon, A. longifolia y A. melanoxylon, respectivamente, (Dennill et al., 1993; Donnelly y Hoffmann, 2004). Esto podría estar relacionado con la baja especificidad de los agentes de control biológico que pueden conducir a afinidades para especies relacionadas (Donnelly y Hoffmann, 2004).
En estos casos, el efecto secundario se puede considerar «positivo» ya que otros congéneres invasivos (todos los que conducen al control de la acacia) fueron atacados. Por lo tanto, el uso de agentes de control biológico en Europa o América del Norte para controlar las acacias debe tener riesgos ecológicos bajos, debido a la ausencia de acacias nativas. Por otro lado, los efectos ecológicos de los agentes introducidos no se abordan completamente y pueden producirse consecuencias inesperadas como reemplazo ecológico, respuestas compensatorias o interacciones entre la red alimentaria (Pearson y Callaway 2003). De hecho, los agentes introducidos en el control de A. longifolia y A. saligna en Sudáfrica crearon redes alimenticias complejas en el área introducida, similares a las de sus rangos nativos (Veldtman et al., 2011).
Recuperación efectiva de áreas limpiadas
La teoría predice que los programas de gestión son más efectivos si los invasores son rápidamente reconocidos y el tiempo entre la introducción y el manejo es lo más corto posible (Simber loff et al., 2015, Kimball y otros, 2015). La intensidad de la intervención necesaria para la recuperación del ecosistema es proporcional a la duración, es decir la densidad, de la invasión (Holmes et al., 2000). Además, la detección temprana de plantas invasoras también contribuye a una gestión rentable. La actual normativa de la Comisión Europea sobre especies invasoras, prevé tres tipos de intervenciones: prevención, detección temprana y erradicación y gestión rápida (EC 2014). Pese a esto, el éxito de la restauración de la tierra después de la eliminación de Acacia es incierto debido a los cambios severos ocurridos en las propiedades físico-químicas del suelo (Marchante et al., 2004, 2011a, b). La transformación de los ecosistemas invadidos por las acacias sugiere que el regreso a condiciones preexistentes es prácticamente imposible; por tanto, el concepto de restauración debe ser entendido como un sinónimo de recuperación.
Después de la eliminación de las acacias invasoras, la recuperación del ecosistema tarda varios años para que los nutrientes y procesos del suelo vuelvan a niveles similares a la pre-invasión. Por tanto, los esfuerzos de erradicación deben mantenerse a largo plazo para lograr resultados positivos (Marchante et al., 2008b).
Para desarrollar programas eficientes de recuperación, también deben considerarse los efectos secundarios después de la eliminación de los invasores. En esta línea, el contenido mejorado de N en suelos invadidos, favorece el asentamiento de hierbas y otros arbustos, pero dificulta las especies ericoides o proteoides (Gaertner et al., 2012). Además, las tasas de crecimiento de la especie nitrofílica Ehrharta calycina aumentaron en los lugares donde se eliminó A. saligna, lo que sugiere que las invasiones subsiguientes por malas hierbas pueden ocurrir después de limpiar las especies exóticas que fijan N2 (Yelenik et al., 2004). Por consiguiente, la recuperación de los ecosistemas puede ser facilitada por la eliminación simultánea de la capa de desechos rica en N, facilitando la germinación de especies nativas a corto plazo (Marchante et al., 2004, 2008b). También es importante considerar que la eliminación de acacias invasoras sin una adecuada planificación de la gestión puede conducir a la exposición del subsuelo infértil vulnerable a la erosión, aún más en las zonas con tasas lentas de colonización de las plantas, como las pendientes de colinas (Van Der Waal et al., 2012). Además, este hecho también restringe la colonización por especies autóctonas que podrían ayudar en la estabilización del suelo (de Neergaard et al., 2005).

Por otro lado, el mantenimiento del banco de semillas nativo es fundamental para recuperar ecosistemas con éxito después de la invasión de Acacia. Se sabe que la recuperación sin éxito de los ecosistemas invadidos después de la eliminación de Acacia está frecuentemente relacionada con la falta de semillas nativas o de suministro de propágulos (Galato witsch y Richardson 2005). Cuando el banco de semillas se agota o alcanza valores críticos, la inclusión de semillas nativas en programas de restauración podría ser esencial para lograr condiciones preexistentes. Además, las especies nativas con bajos requerimientos de nutrientes y una fuerte capacidad competitiva que pueden competir con las acacias invasivas en la etapa temprana de la plántula son particularmente valiosas (Werner et al., 2010), ya que puede facilitar la recuperación del ecosistema.
Al mismo tiempo, los bancos masivos de semillas de acacias son difíciles de gestionar después de la eliminación de las acacias (Richardson y Kluge 2008). En algunos casos, se utilizó fuego para controlarlo en estanques densos invadidos, lo cual reduce el contenido de N en el suelo, provoca la germinación masiva de las semillas de Acacia y ocasionalmente estimula el banco de semillas, como en los ecosistemas propensos a incendios (Le Maitre et al., 2011). Sin embargo, el fuego tiene consecuencias negativas y las quemaduras prescritas sólo se recomiendan en circunstancias específicas, como pendientes pronunciadas o áreas inaccesibles (Fill et al., 2017). En general, el fuego debe ser usado juiciosamente, combinado con otros métodos o descartado en situaciones donde la conservación de la diversidad biológica indígena es de consideración central (Richardson y Kluge 2008). Cabe indicar que la temperatura de la superficie del suelo también puede modificarse sin el uso del fuego. Así en el caso de pequeñas áreas invadidas, la latencia de las semillas de Acacia podría ser eliminada artificialmente mediante la solarización del suelo. Por ejemplo, Cohen et al. (2008) lograron un agotamiento completo de las semillas enterradas de A. saligna utilizando mantillos de polietileno para impedir el proceso fotosintético y producir estrés hidrotérmico.
Por otro lado, hay que indicar que las acciones de restauración activas rara vez se implementan después de limpiar las áreas invadidas, a menos que las relaciones coste / beneficio se consideren aceptables (Fill et al., 2017). La restauración activa puede ser efectiva e incluso financieramente factible, en comparación con la restauración pasiva. De hecho, cada vez hay mayor evidencia de que en algunos casos la restauración de áreas invadidas es factible y puede proporcionar múltiples beneficios sociales y económicos (Murcia et al., 2014).
Hacia una gestión integral de la Acacia en áreas mediterráneas
La experiencia obtenida en la gestión de las acacias ha demostrado que los proyectos exitosos requieren objetivos claros y basados en el tiempo, recursos adecuados y apoyo real y de las partes interesadas (Forsyth et al., 2012). Una estrategia de gestión mejorada debería centrarse en áreas y especies prioritarias, asumiendo los compromisos entre preservar la biodiversidad y evitar la expansión de la invasión, o de lo contrario, el dinero asignado a las acciones de control sería desperdiciado (van Wilgen et al., 2016). La combinación de técnicas de manejo tales como el uso integrado del fuego y la re-siembra activa de áreas despejadas con arbustos indígenas, incrementaría sustancialmente la efectividad de la restauración de ecosistemas (Fill et al., 2017). Sin embargo, hasta ahora, las acciones de manejo realizadas en áreas prioritarias mostraron pocos avances en la reducción de la infestación total (van Wilgen et al., 2012; Gwate et al., 2016).
Hay que destacar que se están desarrollando enfoques de modelación espaciotemporal, tales como modelos basados en individuos, metodología dinámica estocástica o modelos de distribución de especies (SDMs) para anticipar las invasiones de Acacia y gestionar sus impactos en áreas mediterráneas (Thompson et al., 2011; et al., 2015). Recientemente se ha desarrollado un marco jerárquico que combina SDMs, análisis de escenarios y análisis de costes para mejorar la evaluación de invasiones de Acacia a escalas regionales y locales (Vicente et al., 2016). Además de los enfoques anteriores, los impactos de las acacias en un área específica pueden evaluarse inicialmente utilizando el sistema genérico de puntuación de impacto (GISS), una herramienta nueva y factible para cuantificar fácilmente los impactos en los ecosistemas (Nentwig et al., 2016).
Según los autores de este trabajo sobre la invasión de Acacia en áreas mediterráneas, la visión actual de la gestión de acacias por parte de los científicos se centra principalmente en la perspectiva ecológica, evitando las implicaciones socioeconómicas. Generalmente, las acciones de gestión se llevan a cabo con fuentes públicas, resultando en una inevitable necesidad de alianzas sociales y científicas. La percepción pública de los IAP es un elemento clave en la evaluación de las estrategias de gestión, proporcionando un entorno social y político favorable, esencial para lograr resultados exitosos. El uso de las investigaciones está ganando interés como una herramienta informativa y de retroalimentación en los procesos de toma de decisiones (Verbrugge y otros, 2014; Liu y Cook, 2016), ya que de lo contrario, los esfuerzos de erradicación serían inútiles cuando la administración y las acciones sociales no persiguen intereses similares, lo que sugiere que las comunidades locales deben participar activamente en el control del PAI y en los programas de gestión (Mukwada y Manatsa, 2017).
Por otra parte, aspectos socioeconómicos como la migración humana forzada de las zonas rurales a las urbanas conducen al abandono de la tierra y al uso indebido, y este movimiento está favoreciendo la invasión de Acacia y otros IAPs. Por lo que, facilitar el asentamiento de la población en las zonas rurales ayudaría a identificar y evitar rápidamente la dispersión de propágulos de Acacia, preservando la vegetación nativa rural. De hecho, aumentar el acceso al uso de la tierra con fines agrícolas podría resultar en una mayor preocupación, cuidado y, en última instancia, una mejor gestión de las acacias (de Neergaard et al., 2005); sin embargo, desafortunadamente, las tierras no trabajadas o desprotegidas no representan un valor significativo para la sociedad. Para los autores, las políticas gubernamentales centradas exclusivamente en el control de las IAP pero que no evitan el problema del uso indebido de la tierra, no pueden ser totalmente efectivas. De manera idealista, en el contexto actual de un escenario económico cambiante y de un consumo insostenible de recursos, se requieren políticas que adopten iniciativas a largo plazo para mejorar las condiciones de vida humana, reorganizar nuestros conceptos de progreso humano, sociedad sostenible y desarrollo de la tierra.
Discusión y Conclusión
“¿Aquí para quedarse?” fue una pregunta retórica propuesta por Richardson et al. (2011) explorando la dimensión humana-histórica y científico-social de las acacias introducidas. Según la opinión de los autores del presente trabajo, las invasiones de Acacia están lejos de ser plenamente comprendidas y previsibles, convirtiéndose en una tarea desafiante para las próximas décadas. En un contexto de cambio climático y con alteraciones del uso de la tierra, los ecosistemas mediterráneos están bajo presión de nuevas invasiones por especies de Acacia. En este sentido, las herramientas emergentes como el modelado, la genómica, la teledetección y las nuevas herramientas de imagen, la elaboración de bases de datos ecológicas mejoradas o la aplicación y mejora de las ecuaciones alométricas para la estimación de la biomasa a partir de conjuntos de datos forestales más amplios, contribuyen a responder a las preguntas pasadas y futuras con respecto a las invasiones de Acacia. Según su experiencia, las acacias deben considerarse como una entidad en lugar de un grupo de individuos, debido a la reproducción vegetativa masiva. Así, la clonalidad, la integración fisiológica o la asignación de recursos, son temas que siguen siendo poco conocidos para las acacias invasoras.
Hay que tener en cuenta que las acacias se catalogan como plantas indeseables mientras que, al mismo tiempo, su cultivo también proporciona recursos rentables en diferentes países. Por lo tanto, es fundamental determinar el equilibrio entre el valor comercial y los problemas ambientales relacionados. Para evitar los impactos indeseables sin interferir con los propósitos de la industria, se está investigando la implementación de linajes estériles de acacias (Beck y Fossey 2007; Beck-Pay 2013). También sugieren que los administradores forestales, las industrias o los propietarios de tierras que se beneficien del cultivo de acacias exóticas, sean económicamente responsables de los problemas derivados de sus plantaciones. Para evitar la introducción de acacias invasoras es necesario reforzar la ley para unificar las regulaciones forestales, especialmente entre países de la cuenca mediterránea como España, Portugal o Italia.

Las condiciones socioeconómicas actuales son inestables en muchos países, lo que implica que se deben preferir inversiones de gestión rentables en lugar de aquellas que impliquen únicamente costes. En esta línea, se sugiere que la obtención de beneficios a partir de los residuos de la gestión de las acacias podría aliviar el coste de la gestión. Por lo tanto, se ha compilado varias áreas de investigación incipientes donde las acacias podrían ser útiles:
- Agricultura:
Los compuestos de fitotoxicidad de las acacias invasoras podrían utilizarse como base para el desarrollo de nuevos herbicidas biológicos, biopesticidas o mulch fitotóxicos para controlar las malas hierbas o las plagas de los cultivos (Naray, Jabran et al., 2015). De hecho, los estudios para identificar la actividad fitotóxica de compuestos químicos de diferentes materiales de A. dealbata y el uso de abonos verdes de A. dealbata y A. longifolia como bioherbicidas en suelos agrícolas (Souza-Alonso et al en revisión) están actualmente en curso. Al igual que otras especies de leguminosas, las acacias poseen hojas enriquecidas en nutrientes que podrían utilizarse como fertilizantes y una fuente de nutrientes para los cultivos. Además, después de la maduración completa del compost, A. longifolia y A. melanoxylon proporcionan enmiendas agrícolas, biocompuestos, con alto contenido de materia orgánica y baja conductividad eléctrica (Brito et al., 2013, 2015). Los residuos de compost de A. dealbata con lodos de depuradora también mejoran las propiedades bioquímicas y químicas del suelo (Tejada et al., 2014).
- Industria:
El alto contenido de polisacáridos de A. dealbata es un recurso valioso para las biorrefinerías, lo que proporciona una forma de mejorar las materias primas renovables subutilizadas (Yañez et al., 2009, 2013). También se sintetizaron nuevos coagulantes poliméricos catiónicos para el agua y diferentes tipos de tratamiento de efluentes industriales con taninos extraídos de A. mearnsii (Beltrán-Heredia et al., Sánchez-Martín et al.2012, Soares et al., 2012), teniendo también potencial como fitoextratador en la remediación de biosólidos contaminados con metales pesados (Mok et al., 2013). Del mismo modo, Kumari y Ravindhranath (2012) emplearon con éxito A. melanoxylon como biosorbente en la extracción de iones Al + 3 de aguas residuales recogidas de efluentes industriales y lagos contaminados. Además, los extractos de A. mearnsii mostraron resultados positivos para controlar las floraciones de algas azules (Zhou et al., 2012).
- Salud:
Las acacias también pueden ser una fuente química de componentes químicos con fines médicos y de salud. Por ejemplo, la corteza de A. mearnsii se utiliza tradicionalmente en el tratamiento de enfermedades del estómago (Wintola et al., 2 017). Los extractos brutos de esta especie también exhibieron una actividad antimicrobiana significativa, convirtiéndose en una fuente potencial de compuestos bioactivos (Olajuyigbe y Afolayan 2 012). Los contenidos fenólicos, flavonoides y alcaloides de los extractos crudos de A. dealbata y A. melanoxylon mostraron una mayor actividad antioxidante (Luis et al., 2012). Los resultados preliminares también indican que los compuestos solubles en agua presentes en extractos de A. melanoxylon muestran actividad antihelmíntica contra el desarrollo larval de parásitos de caballo (Payne et al., 2013). También la miel de Acacia induce la expresión de citocinas y una metaloproteinasa que degrada el colágeno IV involucrado en la desorganización de la membrana basal durante el proceso de re-epitelización de heridas (Burlando y Cornara, 2013).
- Cosméticos:
Los aceites absolutos de A. dealbata se han utilizado en las industrias cosméticas, especialmente en la producción de perfumes, debido a la presencia de compuestos odorantes (Perriot et al., 2010).
Durante los últimos años se han realizado esfuerzos considerables para abordar las consecuencias de la invasión de Acacia en áreas mediterráneas. Hoy en día, las acacias son reconocidas como amenazas severas para los ecosistemas mediterráneos y el refuerzo de las regulaciones transnacionales, junto con el desarrollo de plataformas de información cruzada, parece crucial para prevenir nuevas introducciones de Acacia. Bajo un escenario futuro de cambio climático, se espera que estos ecosistemas estén en gran parte ocupados por acacias invasoras debido a su mayor crecimiento bajo condiciones más altas de CO2, producción de semilla y resistencia al fuego. Además, los cambios en la dinámica hidrológica por las invasiones de acacias también pueden exacerbar las sequías en las áreas mediterráneas bajo los eventos climáticos extremos esperados.
Las invasiones de las acacias generalmente conducen a cambios en los servicios de los ecosistemas como los regímenes de agua y fuego, la reducción de la biodiversidad de las plantas y la alteración de las propiedades y funciones fisico-químicas del suelo. Las comunidades microbianas modificadas del suelo pueden tener implicaciones negativas para el ciclo de los nutrientes, los procesos de los ecosistemas y la vegetación nativa que dependen de ellos, lo que a su vez podría favorecer la invasión de acacias y aumentar la vulnerabilidad de los ecosistemas afectados. En cuanto a la evaluación de las comunidades de plantas nativas, se debe considerar un conocimiento más profundo de la diversidad funcional y filogenética, en lugar del uso de índices clásicos de diversidad, para evaluar el alcance de los impactos ecológicos producidos. También se necesita trabajo adicional para dilucidar la proporción de reproducción sexual vs. vegetativa durante el proceso de invasión para diseñar estrategias de control adecuadas.
Los autores de este trabajo sobre los conocimientos en la invasión de Acacia en áreas mediterráneas, consideran que la gestión de las acacias debe centrarse en la priorización de la preservación de hábitats no invadidos y la identificación de áreas con potencial para albergar acacias invasivas. También son deseables estudios de evaluación de riesgos basados en marcos más centrados en la predicción y prevención de introducciones futuras, en lugar de evaluar cambios en áreas ya invadidas; así como también es tiempo de comunicarse e involucrar a las percepciones sociales, políticas y las partes interesadas, para proporcionar soluciones integradoras, sostenibles y adaptadas a la invasión de Acacia en áreas mediterráneas; ya que los altos índices económicos no aseguran necesariamente el éxito en el control de las invasiones. Además, hay que destacar que la búsqueda de usos potenciales de los residuos de acacia podría traer soluciones para aliviar parcialmente los recursos económicos asignados a su manejo y, al mismo tiempo, reducir la extensión de las poblaciones invasoras. Por lo tanto, la investigación aplicada sobre usos rentables para residuos de acacia parece ser muy relevante en el futuro.
Finalmente, se concluye que la pregunta retórica planteada hace 6 años “¿aquí para quedarse?” debería estar actualmente transformada -como indica el título original de este trabajo- en una oración afirmativa; ya que la hipótesis emergente de que la erradicación completa de las acacias parece en algunos casos inviable, proporciona un nuevo contexto en el que el estudio del papel ecológico de las formaciones de Acacia -como nuevos ecosistemas- resulta relevante.
Fuente publicación
Este artículo está publicado originalmente en la revista Annals of Forest Sicence, bajo la referencia: Souza-Alonso, P., Rodríguez, J., González, L. et al. Annals of Forest Science (2017) 74: 55. DOI 10.1007/s13595-017-0651-0, estando disponible de forma online para su lectura en la página web de Springer.